Expert médical de l'article

Nouvelles publications
glande pituitaire
Dernière revue: 04.07.2025

Tout le contenu iLive fait l'objet d'un examen médical ou d'une vérification des faits pour assurer autant que possible l'exactitude factuelle.
Nous appliquons des directives strictes en matière d’approvisionnement et ne proposons que des liens vers des sites de médias réputés, des instituts de recherche universitaires et, dans la mesure du possible, des études évaluées par des pairs sur le plan médical. Notez que les nombres entre parenthèses ([1], [2], etc.) sont des liens cliquables vers ces études.
Si vous estimez qu'un contenu quelconque de notre contenu est inexact, obsolète ou discutable, veuillez le sélectionner et appuyer sur Ctrl + Entrée.
L'hypophyse (hypophyse, glandula pituitaria) est située dans la fosse hypophysaire de la selle turcique de l'os sphénoïde et est séparée de la boîte crânienne par un prolongement de la dure-mère cérébrale, formant le diaphragme de la selle. Par l'ouverture de ce diaphragme, l'hypophyse est reliée à l'infundibulum de l'hypothalamus du diencéphale. Ses dimensions transversales sont de 10 à 17 mm, antéropostérieures de 5 à 15 mm et verticales de 5 à 10 mm. Son poids est d'environ 0,5 g chez l'homme et de 0,6 g chez la femme. L'hypophyse est recouverte extérieurement d'une capsule.
Conformément au développement de l'hypophyse à partir de deux rudiments différents, on distingue deux lobes: antérieur et postérieur. L'adénohypophyse, ou lobe antérieur (adénohypophysis, s.lobus anterior), est plus volumineux, représentant 70 à 80 % de la masse totale de l'hypophyse. Il est plus dense que le lobe postérieur. Dans le lobe antérieur, on distingue une partie distale (pars distalis), qui occupe la partie antérieure de la fosse pituitaire, une partie intermédiaire (pars intermedia), située à la limite du lobe postérieur, et une partie tubéreuse (pars tuberalis), s'élevant vers le haut et se connectant à l'infundibulum de l'hypothalamus. En raison de l'abondance de vaisseaux sanguins, le lobe antérieur est de couleur jaune pâle avec une teinte rougeâtre. Le parenchyme de l'hypophyse antérieure est constitué de plusieurs types de cellules glandulaires, entre les faisceaux desquelles se trouvent des capillaires sanguins sinusoïdaux. La moitié (50 %) des cellules adénohypophysaires sont des adénocytes chromophiles, dont le cytoplasme contient des granules fins qui se colorent bien aux sels de chrome. Il s'agit d'adénocytes acidophiles (40 % de l'ensemble des cellules adénohypophysaires) et d'adénocytes basophiles (10 %). Les adénocytes basophiles comprennent les endocrinocytes gonadotropes, corticotropes et thyréotropes. Les adénocytes chromophobes sont petits, possèdent un gros noyau et un faible cytoplasme. Ces cellules sont considérées comme les précurseurs des adénocytes chromophiles. Les 50 % restants sont des adénocytes chromophobes.
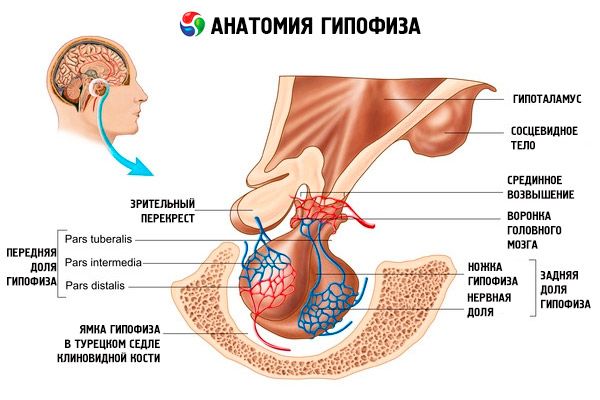
La neurohypophyse, ou lobe postérieur (neurohypophysis, s.lobus posterior), est composée du lobe nerveux (lobus nervosus), situé dans la partie postérieure de la fosse pituitaire, et de l'entonnoir (infundibulum), situé derrière la partie tubéreuse de l'adénohypophyse. Le lobe postérieur de l'hypophyse est formé de cellules neurogliales (cellules hypophysaires), de fibres nerveuses reliant les noyaux neurosécréteurs de l'hypothalamus à la neurohypophyse, et de corpuscules neurosécréteurs.
L'hypophyse, par l'intermédiaire de fibres nerveuses (voies) et de vaisseaux sanguins, est fonctionnellement reliée à l'hypothalamus du diencéphale, qui régule son activité. L'hypophyse et l'hypothalamus, ainsi que leurs connexions neuroendocrines, vasculaires et nerveuses, sont généralement considérés comme le système hypothalamo-hypophysaire.
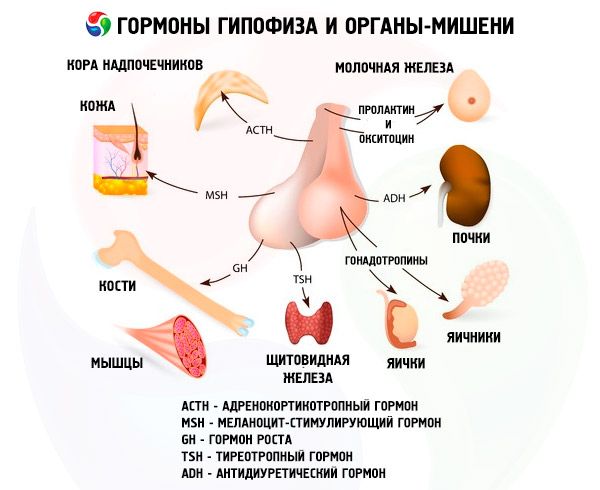
Les hormones de l'hypophyse antérieure et postérieure influencent de nombreuses fonctions de l'organisme, principalement par l'intermédiaire d'autres glandes endocrines. Dans l'hypophyse antérieure, les adénocytes acidophiles (cellules alpha) produisent l'hormone somatotrope (hormone de croissance), qui participe à la régulation de la croissance et du développement des jeunes organismes. Les endocrinocytes corticotropes sécrètent l'hormone adrénocorticotrope (ACTH), qui stimule la sécrétion d'hormones stéroïdes par les glandes surrénales. Les endocrinocytes thyréotropes sécrètent l'hormone thyréotrope (TSH), qui influence le développement de la glande thyroïde et active la production de ses hormones. Les hormones gonadotropes: folliculostimulantes (FSH), lutéinisantes (LH) et prolactine, influencent la maturation sexuelle, régulent et stimulent le développement des follicules ovariens, l'ovulation, la croissance des glandes mammaires et la production de lait chez la femme, ainsi que la spermatogenèse chez l'homme. Ces hormones sont produites par les adénocytes basophiles (cellules bêta). Des facteurs lipotropes de l'hypophyse sont également sécrétés à cet endroit, influençant la mobilisation et l'utilisation des graisses dans l'organisme. Dans la partie intermédiaire du lobe antérieur, se forme l'hormone mélanocytaire stimulante, qui contrôle la formation de pigments – les mélanines – dans l'organisme.
Les cellules neurosécrétrices des noyaux supraoptique et paraventriculaire de l'hypothalamus produisent de la vasopressine et de l'ocytocine. Ces hormones sont transportées vers les cellules de la posthypophyse par les axones qui constituent le tractus hypothalamo-hypophysaire. De la posthypophyse, ces substances passent dans la circulation sanguine. La vasopressine a un effet vasoconstricteur et antidiurétique, ce qui lui vaut son nom d'hormone antidiurétique (ADH). L'ocytocine stimule la contractilité des muscles utérins, augmente la sécrétion de lait par la glande mammaire en lactation, inhibe le développement et le fonctionnement du corps jaune et influence le tonus des muscles lisses (non striés) du tube digestif.
Développement de l'hypophyse
Le lobe antérieur de l'hypophyse se développe à partir de l'épithélium de la paroi dorsale de la cavité buccale, formant une excroissance annulaire (poche de Rathke). Cette protubérance ectodermique se développe vers la base du futur troisième ventricule. Vers elle, à partir de la face inférieure de la deuxième vésicule cérébrale (futur fond du troisième ventricule), se développe un prolongement duquel se développent le tubercule gris de l'infundibulum et le lobe postérieur de l'hypophyse.
Vaisseaux et nerfs de l'hypophyse
Depuis les artères carotides internes et les vaisseaux du cercle artériel cérébral, les artères hypophysaires supérieure et inférieure sont dirigées vers l'hypophyse. Les artères hypophysaires supérieures rejoignent le noyau gris et l'infundibulum de l'hypothalamus, où elles s'anastomosent et forment des capillaires pénétrant le tissu cérébral: le réseau hémocapillaire primaire. Les anses longues et courtes de ce réseau forment les veines portes, dirigées vers le lobe antérieur de l'hypophyse. Dans le parenchyme du lobe antérieur de l'hypophyse, ces veines se divisent en larges capillaires sinusoïdaux, formant le réseau hémocapillaire secondaire. Le lobe postérieur de l'hypophyse est irrigué principalement par l'artère hypophysaire inférieure. De longues anastomoses artérielles existent entre les artères hypophysaires supérieure et inférieure. L'écoulement du sang veineux du réseau hémocapillaire secondaire s'effectue par un système de veines qui se jettent dans les sinus caverneux et intercaverneux de la dure-mère du cerveau.
Les fibres sympathiques qui pénètrent dans l'organe avec les artères participent à l'innervation de l'hypophyse. Les fibres nerveuses sympathiques postganglionnaires partent du plexus de l'artère carotide interne. De plus, de nombreuses terminaisons des prolongements des cellules neurosécrétoires situées dans les noyaux de l'hypothalamus se trouvent dans le lobe postérieur de l'hypophyse.
Caractéristiques de l'hypophyse liées à l'âge
Le poids moyen de l'hypophyse chez le nouveau-né atteint 0,12 g. Ce poids double vers 10 ans et triple vers 15 ans. À 20 ans, le poids de l'hypophyse atteint son maximum (530-560 mg) et reste quasiment inchangé par la suite. Après 60 ans, on observe une légère diminution du poids de cette glande endocrine.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hormones hypophysaires
L'unité de la régulation nerveuse et hormonale de l'organisme est assurée par l'étroite connexion anatomique et fonctionnelle de l'hypophyse et de l'hypothalamus. Ce complexe détermine l'état et le fonctionnement de l'ensemble du système endocrinien.
L'hypophyse est la principale glande endocrine produisant un certain nombre d'hormones peptidiques régulant directement la fonction des glandes périphériques. Il s'agit d'une formation en forme de haricot gris-rouge, recouverte d'une capsule fibreuse pesant de 0,5 à 0,6 g. Sa taille varie légèrement selon le sexe et l'âge. La division généralement admise de l'hypophyse en deux lobes, distincts par leur développement, leur structure et leur fonction, demeure: l'adénohypophyse distale antérieure et l'hypophyse postérieure. Le premier, qui représente environ 70 % de la masse totale de la glande, est classiquement divisé en parties distale, infundibulaire et intermédiaire, tandis que le second est divisé en partie postérieure, ou lobe, et en tige pituitaire. La glande est située dans la fosse pituitaire de la selle turcique de l'os sphénoïde et est reliée au cerveau par cette tige. La partie supérieure du lobe antérieur est recouverte par le chiasma optique et les tractus optiques. L'hypophyse est très abondamment irriguée par les branches de l'artère carotide interne (artères hypophysaires supérieure et inférieure), ainsi que par celles du cercle artériel cérébral. Les artères hypophysaires supérieures participent à l'irrigation sanguine de l'adénohypophyse, tandis que les artères hypophysaires inférieures participent à l'irrigation sanguine de la neurohypophyse, en contact avec les terminaisons neurosécrétoires des axones des noyaux à grandes cellules de l'hypothalamus. Les premières pénètrent dans l'éminence médiane de l'hypothalamus, où elles se dispersent dans un réseau capillaire (le plexus capillaire primaire). Ces capillaires (avec lesquels entrent en contact les terminaisons des axones des petites cellules neurosécrétoires de l'hypothalamus médiobasal) se rassemblent dans les veines portes qui descendent le long de la tige pituitaire jusqu'au parenchyme de l'adénohypophyse, où elles se divisent à nouveau en un réseau de capillaires sinusoïdaux (le plexus capillaire secondaire). Ainsi, le sang, après avoir traversé au préalable l'éminence médiane de l'hypothalamus, où il s'enrichit en hormones adénohypophysotropes hypothalamiques (hormones de libération), pénètre dans l'adénohypophyse.
L'écoulement du sang saturé d'hormones adénohypophysaires depuis les nombreux capillaires du plexus secondaire s'effectue par le réseau veineux, qui à son tour s'écoule dans les sinus veineux de la dure-mère, puis dans la circulation sanguine générale. Ainsi, le système porte de l'hypophyse, avec un flux sanguin descendant depuis l'hypothalamus, est une composante morphofonctionnelle du mécanisme complexe de contrôle neurohumoral des fonctions tropiques de l'adénohypophyse.
L'hypophyse est innervée par des fibres sympathiques qui suivent les artères hypophysaires. Elles proviennent des fibres postganglionnaires qui traversent le plexus carotidien interne, connecté aux ganglions cervicaux supérieurs. L'adénohypophyse n'est pas innervée directement par l'hypothalamus. Le lobe postérieur reçoit des fibres nerveuses provenant des noyaux neurosécréteurs de l'hypothalamus.
L'adénohypophyse est une formation très complexe dans son architecture histologique. Elle comporte deux types de cellules glandulaires: chromophobes et chromophiles. Ces dernières sont à leur tour divisées en cellules acidophiles et basophiles (une description histologique détaillée de l'hypophyse est donnée dans la section correspondante du manuel). Cependant, il convient de noter que les hormones produites par les cellules glandulaires qui composent le parenchyme de l'adénohypophyse, en raison de la diversité de ces dernières, présentent des caractéristiques chimiques légèrement différentes, et la structure fine des cellules sécrétrices doit correspondre aux caractéristiques de biosynthèse de chacune d'elles. Cependant, des formes transitoires de cellules glandulaires capables de produire plusieurs hormones peuvent parfois être observées dans l'adénohypophyse. Il est prouvé que le type de cellules glandulaires de l'adénohypophyse n'est pas toujours déterminé génétiquement.
Sous le diaphragme de la selle turcique se trouve la partie en forme d'entonnoir du lobe antérieur. Elle entoure la tige pituitaire et est en contact avec le tubercule gris. Cette partie de l'adénohypophyse se caractérise par la présence de cellules épithéliales et une vascularisation abondante. Elle est également hormonalement active.
La partie intermédiaire (moyenne) de l'hypophyse est constituée de plusieurs couches de grandes cellules basophiles sécrétoires actives.
L'hypophyse remplit diverses fonctions grâce à ses hormones. Son lobe antérieur produit des hormones adrénocorticotropes (ACTH), thyréostimulines (TSH), folliculostimulantes (FSH), lutéinisantes (LH), lipotropes, ainsi que l'hormone de croissance somatotrope (STO) et la prolactine. Le lobe intermédiaire synthétise l'hormone mélanocytaire (MSH), tandis que le lobe postérieur accumule la vasopressine et l'ocytocine.
ACTH
Les hormones hypophysaires sont un groupe d'hormones protéiques, peptidiques et glycoprotéines. Parmi les hormones de l'hypophyse antérieure, l'ACTH est celle qui a été la mieux étudiée. Elle est produite par les cellules basophiles. Sa principale fonction physiologique est de stimuler la biosynthèse et la sécrétion d'hormones stéroïdes par le cortex surrénalien. L'ACTH présente également une activité mélanocytaire et lipotrope. Isolée pure en 1953, elle a ensuite été établie. Sa structure chimique, composée de 39 résidus d'acides aminés, a été établie chez l'homme et chez plusieurs mammifères. L'ACTH n'a pas de spécificité d'espèce. Actuellement, la synthèse chimique de l'hormone elle-même et de divers fragments de sa molécule, plus actifs que les hormones naturelles, a été réalisée. La structure de l'hormone comporte deux segments de la chaîne peptidique: l'un assure la détection et la liaison de l'ACTH au récepteur, l'autre a un effet biologique. Il semble que l'ACTH se lie au récepteur de l'ACTH par l'interaction des charges électriques de l'hormone et du récepteur. Le rôle de l'effecteur biologique de l'ACTH est assuré par le fragment de la molécule 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
L'activité mélanocytaire de l'ACTH est due à la présence dans la molécule d'une région N-terminale, composée de 13 acides aminés et reproduisant la structure de l'hormone alpha-mélanocytaire. Cette région contient également un heptapeptide présent dans d'autres hormones hypophysaires et possédant des activités adrénocorticotrope, mélanocytaire et lipotrope.
Le moment clé de l'action de l'ACTH est considéré comme l'activation de l'enzyme protéine kinase dans le cytoplasme avec la participation de l'AMPc. La protéine kinase phosphorylée active l'enzyme estérase, qui convertit les esters de cholestérol en substance libre dans les gouttelettes lipidiques. La protéine synthétisée dans le cytoplasme par phosphorylation des ribosomes stimule la liaison du cholestérol libre au cytochrome P-450 et son transfert des gouttelettes lipidiques vers les mitochondries, où sont présentes toutes les enzymes assurant la conversion du cholestérol en corticostéroïdes.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Hormone stimulant la thyroïde
La TSH (thyrotropine) est le principal régulateur du développement et du fonctionnement de la glande thyroïde, ainsi que des processus de synthèse et de sécrétion des hormones thyroïdiennes. Cette protéine complexe, la glycoprotéine, est constituée de sous-unités alpha et bêta. La structure de la première sous-unité coïncide avec celle de la sous-unité alpha de l'hormone lutéinisante. De plus, elle est largement identique chez différentes espèces animales. La séquence des acides aminés de la sous-unité bêta de la TSH humaine a été déchiffrée et comprend 119 acides aminés. Il est à noter que les sous-unités bêta de la TSH humaine et bovine présentent de nombreuses similitudes. Les propriétés biologiques et la nature de l'activité biologique des hormones glycoprotéiques sont déterminées par la sous-unité bêta. Elle assure également l'interaction de l'hormone avec les récepteurs de divers organes cibles. Cependant, chez la plupart des animaux, la sous-unité bêta ne présente une activité spécifique qu'après s'être combinée à la sous-unité alpha, qui agit comme une sorte d'activateur de l'hormone. Cette dernière induit, avec la même probabilité, des activités lutéinisantes, folliculostimulantes et thyréotropes, déterminées par les propriétés de la sous-unité bêta. La similitude constatée permet de conclure que ces hormones sont issues de l'évolution d'un prédécesseur commun; la sous-unité bêta détermine également leurs propriétés immunologiques. On suppose que la sous-unité alpha protège la sous-unité bêta de l'action des enzymes protéolytiques et facilite son transport de l'hypophyse vers les organes cibles périphériques.
Hormones gonadotropes
Les gonadotrophines sont présentes dans l'organisme sous forme de LH et de FSH. Leur fonction se limite généralement à assurer la reproduction chez les individus des deux sexes. Comme la TSH, elles sont des protéines complexes: des glycoprotéines. La FSH induit la maturation des follicules ovariens chez la femme et stimule la spermatogenèse chez l'homme. La LH provoque la rupture folliculaire chez la femme, entraînant la formation du corps jaune, et stimule la sécrétion d'œstrogènes et de progestérone. Chez l'homme, cette même hormone accélère le développement du tissu interstitiel et la sécrétion d'androgènes. Les effets des gonadotrophines sont interdépendants et synchrones.
La dynamique de la sécrétion de gonadotrophines chez la femme évolue au cours du cycle menstruel et a été étudiée en détail. Durant la phase préovulatoire (folliculaire) du cycle, la concentration de LH est relativement faible, tandis que celle de FSH est augmentée. À mesure que le follicule mûrit, la sécrétion d'œstradiol augmente, ce qui favorise une production accrue de gonadotrophines par l'hypophyse et la survenue de cycles de LH et de FSH. Autrement dit, les stéroïdes sexuels stimulent la sécrétion de gonadotrophines.
Actuellement, la structure de la LH a été déterminée. Comme la TSH, elle est constituée de deux sous-unités: a et b. La structure de la sous-unité alpha de la LH chez différentes espèces animales coïncide largement et correspond à celle de la TSH.
La structure de la sous-unité bêta de la LH diffère sensiblement de celle de la sous-unité bêta de la TSH, bien qu'elle comporte quatre segments identiques de la chaîne peptidique, composés de 4 à 5 résidus d'acides aminés. Dans la TSH, ils sont localisés aux positions 27-31, 51-54, 65-68 et 78-83. Puisque la sous-unité bêta de la LH et de la TSH détermine l'activité biologique spécifique des hormones, on peut supposer que des segments homologues dans la structure de la LH et de la TSH devraient assurer la connexion des sous-unités bêta à la sous-unité alpha, et que les segments de structure différente devraient être responsables de la spécificité de l'activité biologique des hormones.
La LH native est très stable à l'action des enzymes protéolytiques, mais la sous-unité bêta est rapidement clivée par la chymotrypsine, et la sous-unité a est difficile à hydrolyser par l'enzyme, c'est-à-dire qu'elle joue un rôle protecteur, empêchant la chymotrypsine d'accéder aux liaisons peptidiques.
Quant à la structure chimique de la FSH, les chercheurs n'ont pas encore obtenu de résultats définitifs. Comme la LH, la FSH est composée de deux sous-unités, mais la sous-unité bêta de la FSH diffère de celle de la LH.
Prolactine
Une autre hormone, la prolactine (hormone lactogène), joue un rôle actif dans les processus de reproduction. Chez les mammifères, ses principales propriétés physiologiques se manifestent par la stimulation du développement des glandes mammaires et de la lactation, ainsi que par la croissance des glandes sébacées et des organes internes. Elle favorise l'effet des stéroïdes sur les caractères sexuels secondaires chez les mâles, stimule l'activité sécrétoire du corps jaune chez la souris et le rat, et participe à la régulation du métabolisme des graisses. Ces dernières années, la prolactine a fait l'objet d'une attention particulière en tant que régulateur du comportement maternel; cette polyvalence s'explique par son évolution. C'est l'une des plus anciennes hormones hypophysaires, présente même chez les amphibiens. À l'heure actuelle, la structure de la prolactine chez certaines espèces de mammifères est parfaitement élucidée. Cependant, jusqu'à récemment, les scientifiques doutaient de l'existence d'une telle hormone chez l'homme. Nombreux étaient ceux qui pensaient que sa fonction était assurée par l'hormone de croissance. Des preuves convaincantes de la présence de prolactine chez l'homme ont été obtenues et sa structure a été partiellement déchiffrée. Les récepteurs de la prolactine se lient activement à l'hormone de croissance et au lactogène placentaire, indiquant un mécanisme d'action unique des trois hormones.
Somatotropine
L'hormone de croissance, la somatotropine, a un spectre d'action encore plus large que la prolactine. Comme la prolactine, elle est produite par les cellules acidophiles de l'adénohypophyse. La somatotropine stimule la croissance squelettique, active la biosynthèse des protéines, a un effet mobilisateur de graisses et favorise la prise de masse corporelle. De plus, elle coordonne les processus métaboliques.
La participation de l'hormone à ce dernier est confirmée par le fait d'une forte augmentation de sa sécrétion par l'hypophyse, par exemple, lorsque le taux de sucre dans le sang diminue.
La structure chimique de cette hormone humaine est désormais parfaitement établie: elle compte 191 résidus d’acides aminés. Sa structure primaire est similaire à celle de la somatomammotropine chorionique ou du lactogène placentaire. Ces données indiquent une proximité évolutive significative entre les deux hormones, bien qu’elles présentent des différences d’activité biologique.
Il est important de souligner la grande spécificité spécifique de l'hormone en question. Par exemple, la STH d'origine animale est inactive chez l'homme. Cela s'explique à la fois par la réaction entre les récepteurs de la STH chez l'homme et l'animal, et par la structure même de l'hormone. Des études sont actuellement en cours pour identifier les centres actifs de la structure complexe de la STH présentant une activité biologique. Des fragments individuels de la molécule présentant d'autres propriétés sont étudiés. Par exemple, après hydrolyse de la STH humaine par la pepsine, un peptide composé de 14 résidus d'acides aminés et correspondant à la section 31-44 de la molécule a été isolé. Il n'a pas eu d'effet sur la croissance, mais son activité lipotrope a été significativement supérieure à celle de l'hormone native. L'hormone de croissance humaine, contrairement à son homologue animale, possède une activité lactogène significative.
L'adénohypophyse synthétise de nombreuses substances peptidiques et protéiques à effet mobilisateur de graisses, et les hormones hypophysaires tropiques – ACTH, STH, TSH et autres – ont un effet lipotrope. Ces dernières années, les hormones bêta- et y-lipotropes (LPG) ont été particulièrement mises en avant. Les propriétés biologiques de la bêta-LPG ont été étudiées en détail; outre son activité lipotrope, elle a également un effet mélanocytaire, corticotrope et hypocalcémiant, ainsi qu'un effet insulino-similaire.
À l'heure actuelle, la structure primaire de la LPG ovine (90 résidus d'acides aminés), hormones lipotropes porcines et bovines, a été déchiffrée. Cette hormone présente une spécificité d'espèce, bien que la structure de sa région centrale soit identique d'une espèce à l'autre. Elle détermine ses propriétés biologiques. Un fragment de cette région se retrouve dans la structure de l'alpha-MSH, de la bêta-MSH, de l'ACTH et de la bêta-LPG. Il est suggéré que ces hormones sont issues du même précurseur au cours de l'évolution. La γ-LPG présente une activité lipotrope plus faible que la bêta-LPG.
Hormone stimulant les mélanocytes
Cette hormone, synthétisée dans le lobe intermédiaire de l'hypophyse, stimule la biosynthèse de la mélanine, un pigment cutané, et favorise l'augmentation de la taille et du nombre de mélanocytes dans la peau des amphibiens. Ces propriétés de la MSH sont utilisées dans les tests biologiques de l'hormone. Il existe deux types d'hormones: l'alpha-MSH et la bêta-MSH. Il a été démontré que l'alpha-MSH n'est pas spécifique à l'espèce et possède la même structure chimique chez tous les mammifères. Sa molécule est une chaîne peptidique composée de 13 résidus d'acides aminés. La bêta-MSH, en revanche, est spécifique à l'espèce et sa structure diffère selon les animaux. Chez la plupart des mammifères, la molécule de bêta-MSH est composée de 18 résidus d'acides aminés, et ce n'est que chez l'homme qu'elle est prolongée de quatre résidus d'acides aminés à partir de l'extrémité de l'acide aminé. Il convient de noter que l'alpha-MSH possède une certaine activité adrénocorticotrope et que son effet sur le comportement des animaux et des humains est désormais prouvé.
L'ocytocine et la vasopressine
La vasopressine et l'ocytocine, synthétisées dans l'hypothalamus, s'accumulent dans le lobe postérieur de l'hypophyse: la vasopressine dans les neurones du noyau supraoptique et l'ocytocine dans le noyau paraventriculatoire. Elles sont ensuite transférées vers l'hypophyse. Il convient de souligner que le précurseur de l'hormone vasopressine est d'abord synthétisé dans l'hypothalamus. Parallèlement, les protéines neurophysines de types 1 et 2 y sont produites. La première se lie à l'ocytocine, et la seconde à la vasopressine. Ces complexes migrent sous forme de granules neurosécrétoires dans le cytoplasme le long de l'axone et atteignent le lobe postérieur de l'hypophyse, où les fibres nerveuses se terminent dans la paroi vasculaire et où le contenu des granules pénètre dans le sang. La vasopressine et l'ocytocine sont les premières hormones hypophysaires dont la séquence d'acides aminés est parfaitement établie. Leur structure chimique est celle de nonapeptides possédant un pont disulfure.
Les hormones étudiées produisent divers effets biologiques: elles stimulent le transport de l'eau et des sels à travers les membranes, ont un effet vasopresseur, augmentent les contractions des muscles lisses de l'utérus lors de l'accouchement et augmentent la sécrétion des glandes mammaires. Il convient de noter que la vasopressine a une activité antidiurétique supérieure à celle de l'ocytocine, tandis que cette dernière a un effet plus marqué sur l'utérus et les glandes mammaires. Le principal régulateur de la sécrétion de vasopressine est la consommation d'eau; dans les tubules rénaux, elle se lie aux récepteurs des membranes cytoplasmiques, ce qui entraîne l'activation de l'enzyme adénylate cyclase. Différentes parties de la molécule sont responsables de la liaison de l'hormone au récepteur et de l'effet biologique.
L'hypophyse, reliée à l'ensemble du système nerveux par l'hypothalamus, unit le système endocrinien en un tout fonctionnel, contribuant à la stabilité du milieu interne de l'organisme (homéostasie). Au sein du système endocrinien, la régulation homéostatique repose sur le principe de rétroaction entre le lobe antérieur de l'hypophyse et les glandes cibles (thyroïde, cortex surrénal, gonades). Un excès d'hormone produite par la glande cible inhibe, tandis qu'une carence stimule la sécrétion et la libération de l'hormone tropique correspondante. L'hypothalamus fait partie de ce système de rétroaction. C'est en lui que se situent les zones réceptrices sensibles aux hormones des glandes cibles. En se liant spécifiquement aux hormones circulantes et en modifiant leur réponse en fonction de leur concentration, les récepteurs hypothalamiques transmettent leur effet aux centres hypothalamiques correspondants, qui coordonnent le travail de l'adénohypophyse, libérant ainsi les hormones adénohypophysotropes hypothalamiques. Ainsi, l’hypothalamus doit être considéré comme un cerveau neuroendocrinien.